在此页面上
双特异性抗体(bsAbs)通过基因重组、化学偶联或四杂交瘤技术设计和制造,能够在一个抗体分子中同时包含两个靶向结合单元,分别识别两个独立的表位或抗原。通过顺序或同步结合,bsAbs在体内发挥多种作用机制(MoA),作为两个抗原之间的生物物理桥梁,实现特定的治疗效果,包括T细胞和自然杀伤细胞(NK细胞)的招募、肿瘤靶向以及免疫调节。尤其是T细胞介导的双特异性抗体,在类风湿关节炎、系统性红斑狼疮等自身免疫疾病中,通过靶向特定免疫细胞如B细胞和炎症介质,展现出良好的治疗潜力。
目前,许多bsAbs正处于临床试验阶段,主要用于治疗各种疾病,尤其是肿瘤。在此,我们提供科学的动物模型和系统性的生物检测分析方法,支持抗肿瘤双特异性抗体的临床前研究,包括靶点验证、安全性评估和药效评价等方面。
百奥赛图建立了先进的药理药效平台,支持新型抗体的应用开发。针对双特异性抗体的临床前研究,我们开发了多种动物模型以满足定制化需求,涵盖同种移植模型、异种移植模型、靶点人源化小鼠模型(如B-hCD3E和B-hCD3EDG小鼠)及人免疫系统重建模型。在体外研究方面,我们的流式细胞术平台和细胞平台能够快速、可靠且可重复地开展一系列功能性和机制研究,包括靶点验证、细胞内信号转导、抗体Fc功能(ADCC、ADCP、CDC)及安全性评估等,为双特异性抗体的研发提供坚实支持。
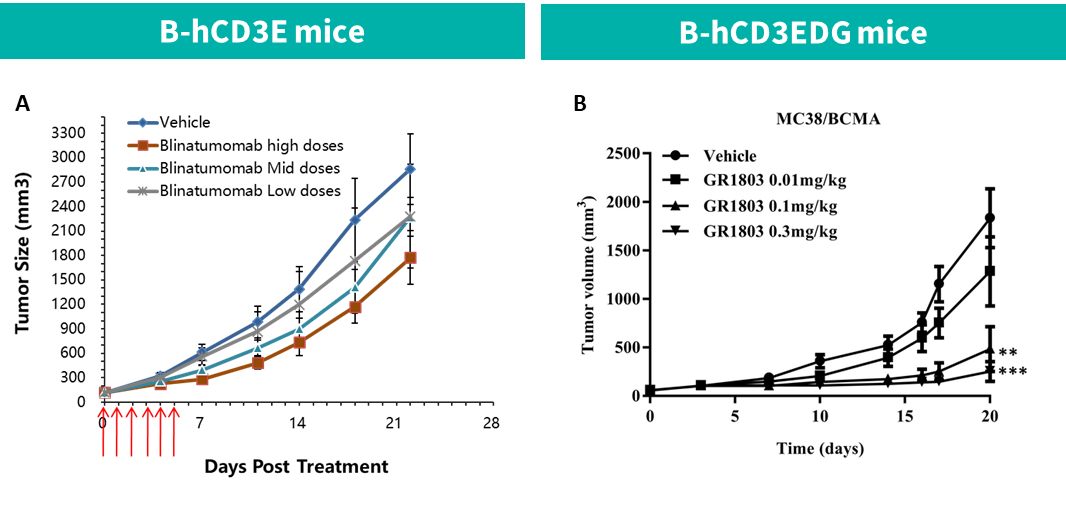
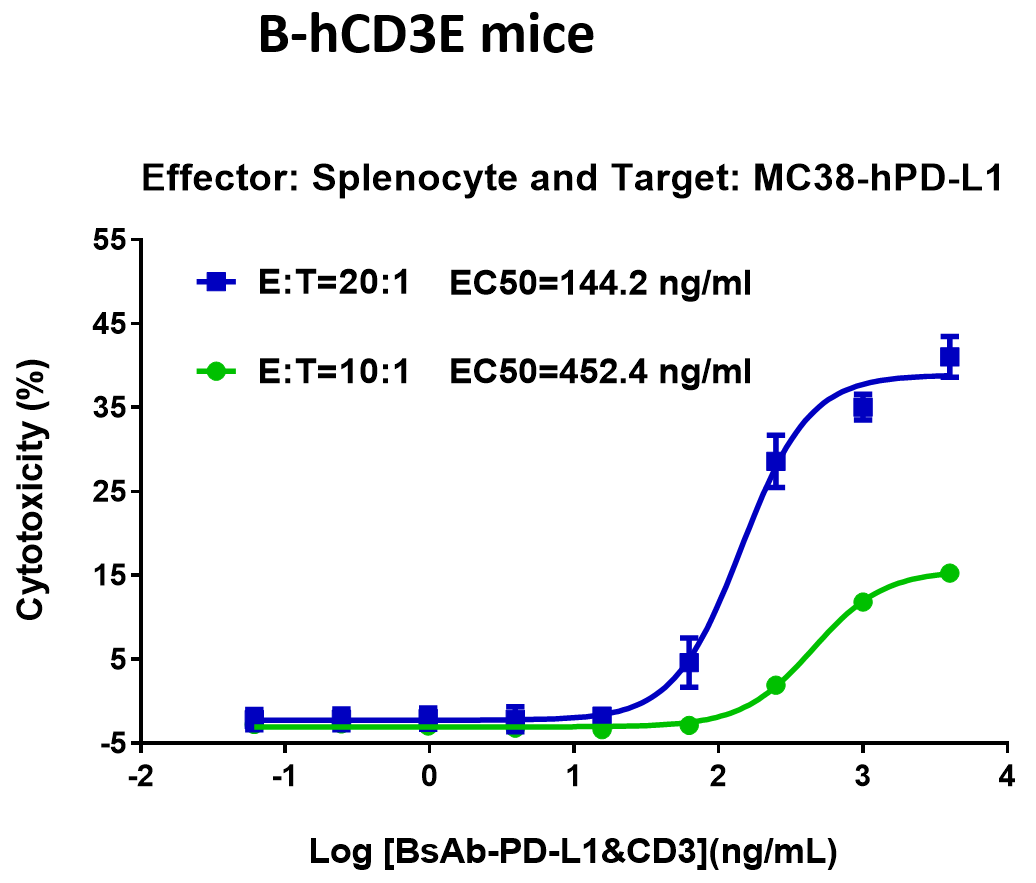
(A) Blinatumomab在B-hCD3E小鼠中的抗肿瘤活性。
B-hCD19 MC38细胞被皮下植入B-hCD3E小鼠。当肿瘤大小约为50-100mm3时,将小鼠分组。
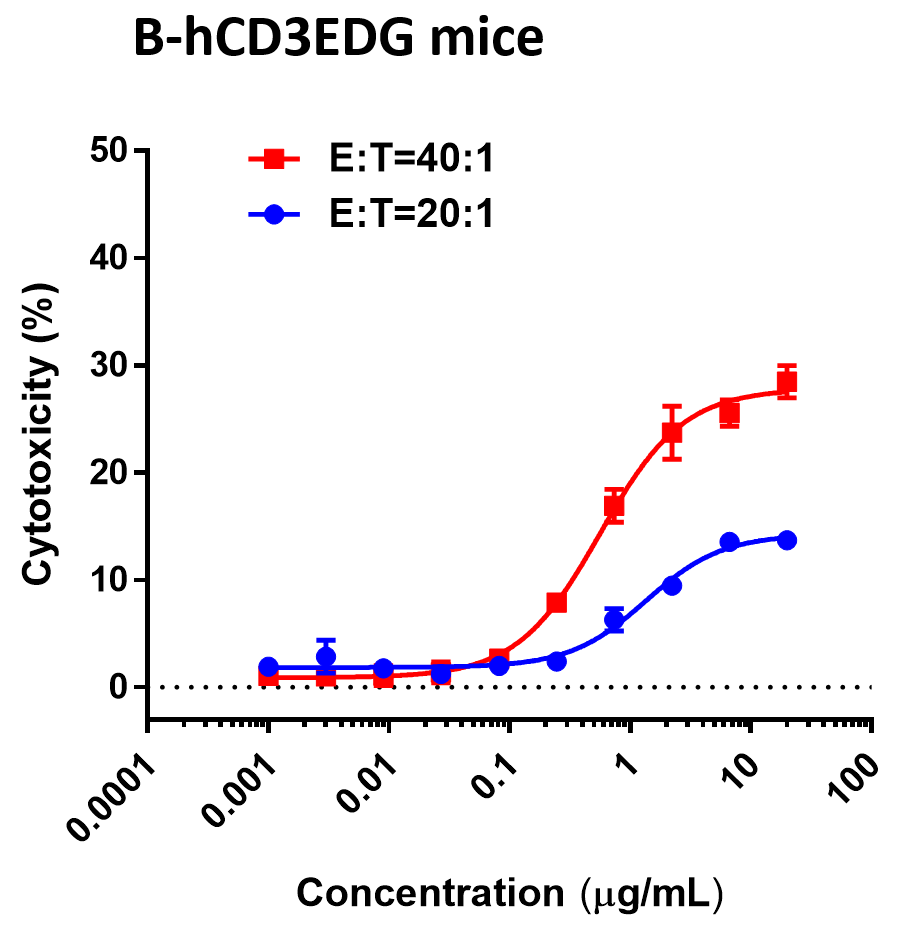
(B) 抗体GR1803在B-hCD3EDG小鼠中的抗肿瘤活性。
B-hBCMA MC38细胞被皮下植入B-hCD3EDG小鼠(雌性,6-8周龄)。当肿瘤大小约为50-100mm3时,将小鼠分组。(Li, L., Chen, X., Li, Y., & Lou, L. (2022).)
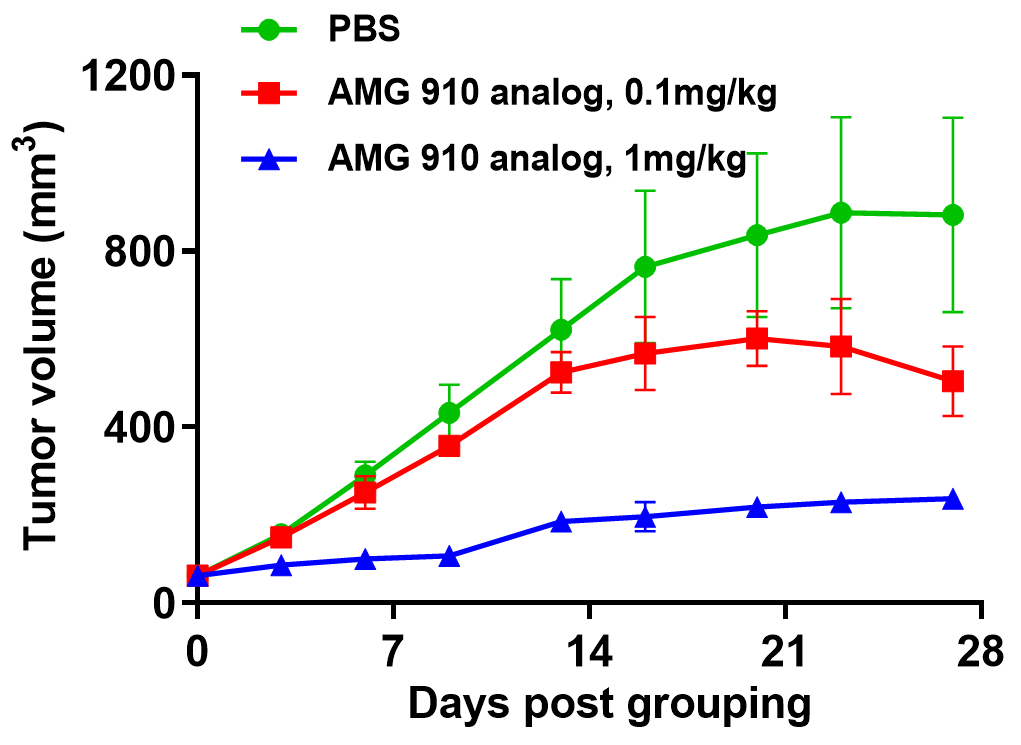
双特异性抗体在人免疫系统重建模型中的药效评估。 (A) CD3双特异性抗体研究的人免疫系统重建模型。将NUGC4细胞(5E6)皮下植入B-NDG小鼠,在将人PBMC(5E6)静脉注射后,抗人CD3×Claudin18.2双特异性抗体(AMG 910类似物)显著抑制了人PBMC重建的B-NDG小鼠中NUGC4肿瘤的生长。
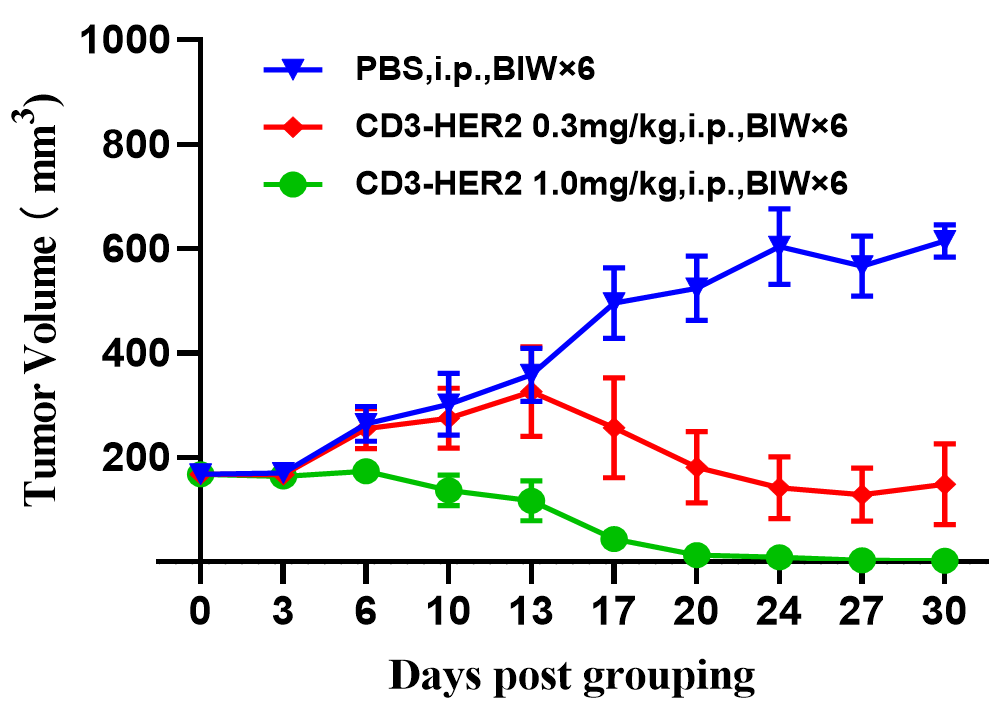
人PBMC重建B-NDG小鼠用于CD3×HER2双特异性抗体的体内药效研究。 NCI-N87细胞(1E7)在静脉注射人PBMC(5E6)3天前皮下植入B-NDG小鼠。当肿瘤大小约为100-150mm3,且人血液中hCD45%的百分比超过10%时,按对照组和治疗组分组进行治疗。结果显示,抗人CD3×HER2双特异性抗体显著抑制了在人PBMC重建的B-NDG小鼠模型中中NCI-N87肿瘤的生长。
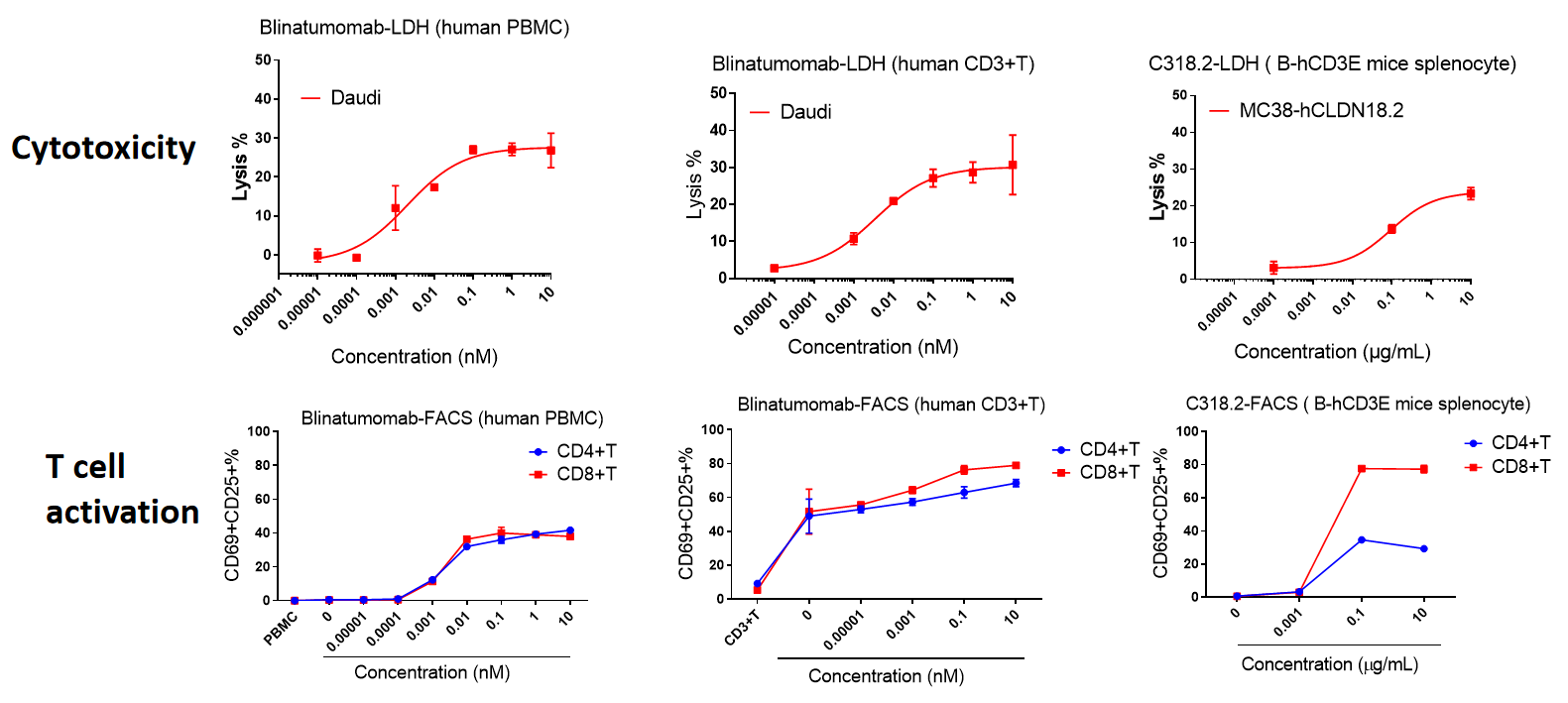
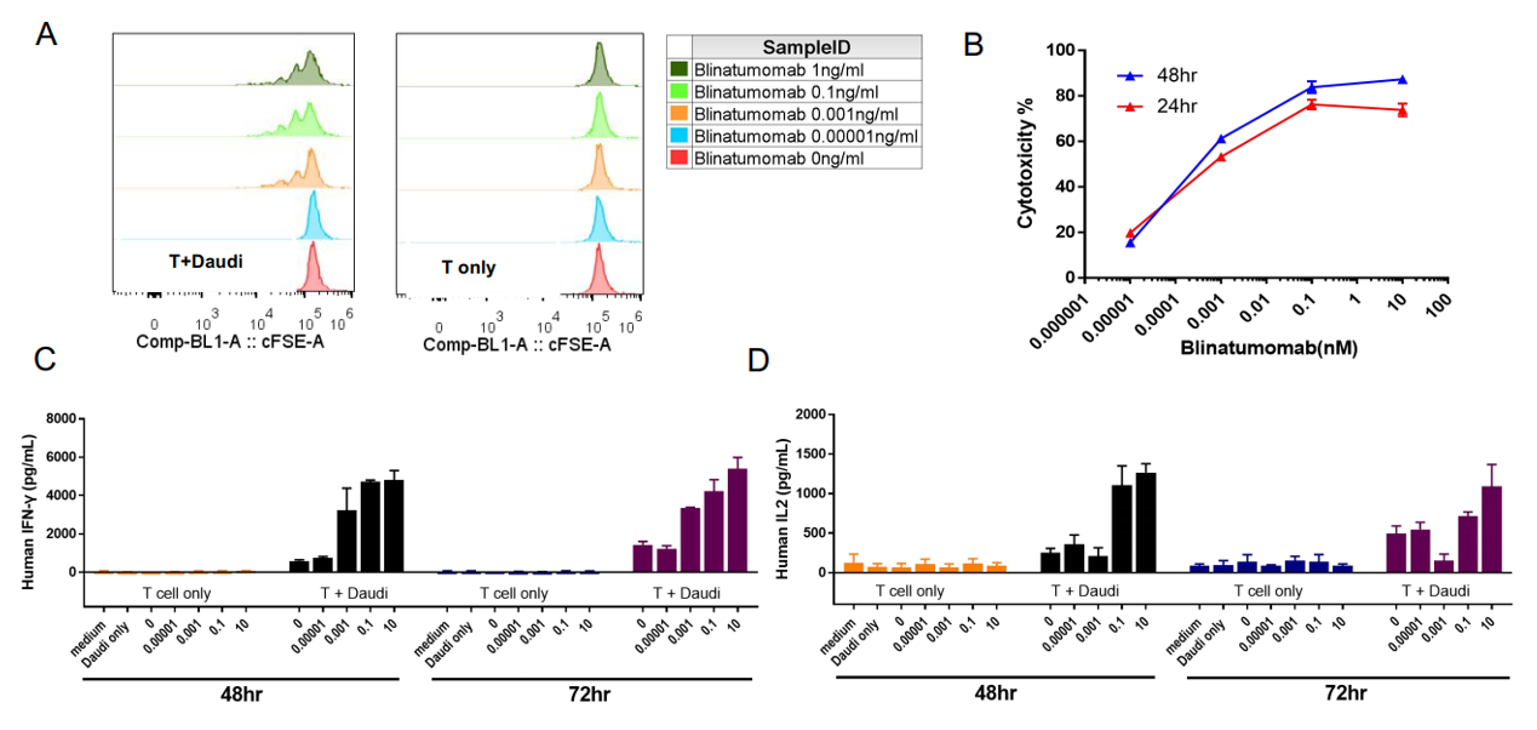
A: T细胞与Daudi细胞共培养时,Blinatumomab以剂量依赖的方式促进T细胞的增殖。B: Blinatumomab的细胞毒性以剂量依赖的方式增强(Daudi与T细胞的比例为10:1)。C-D: Blinatumomab以剂量依赖的方式促进IFN-γ和IL-2的分泌。
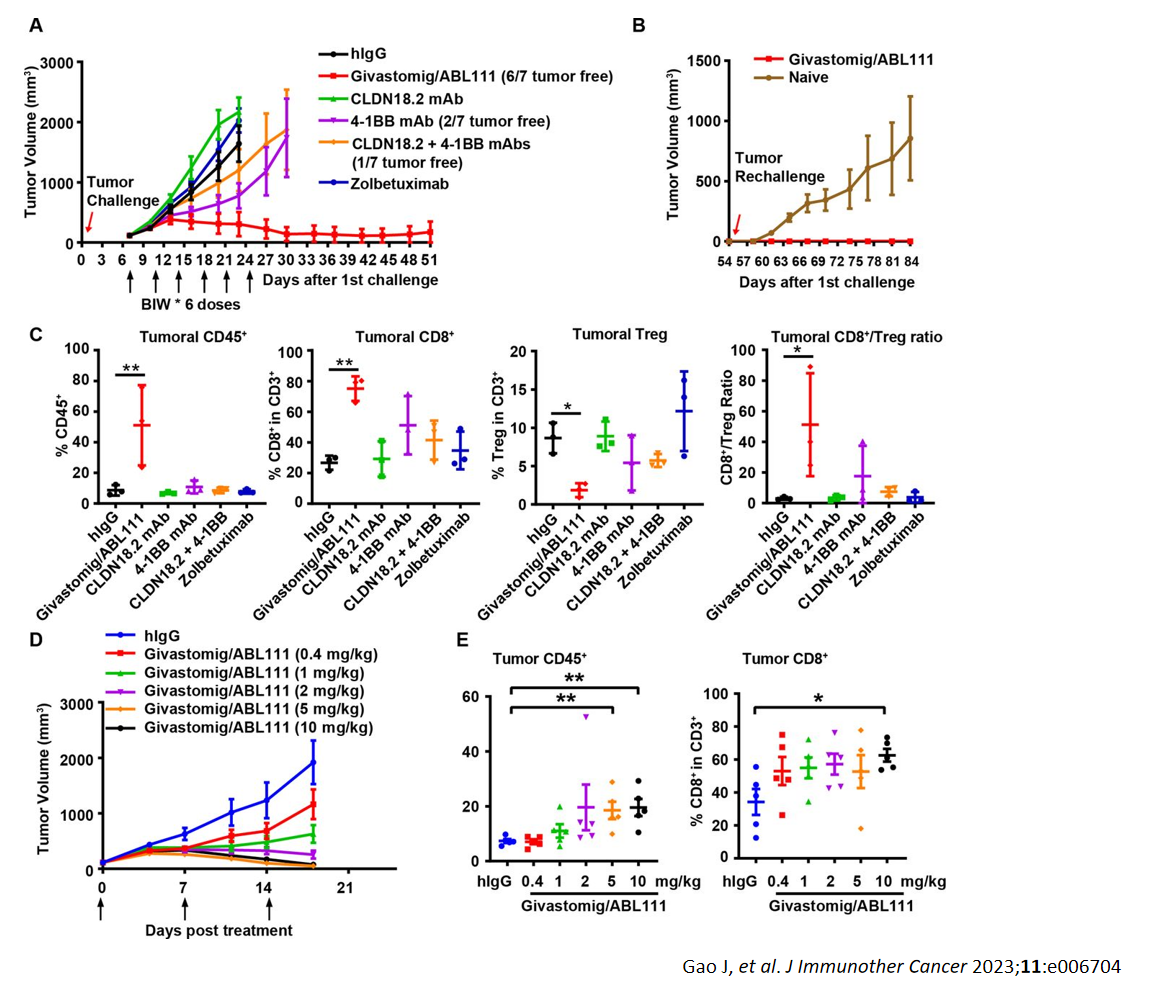
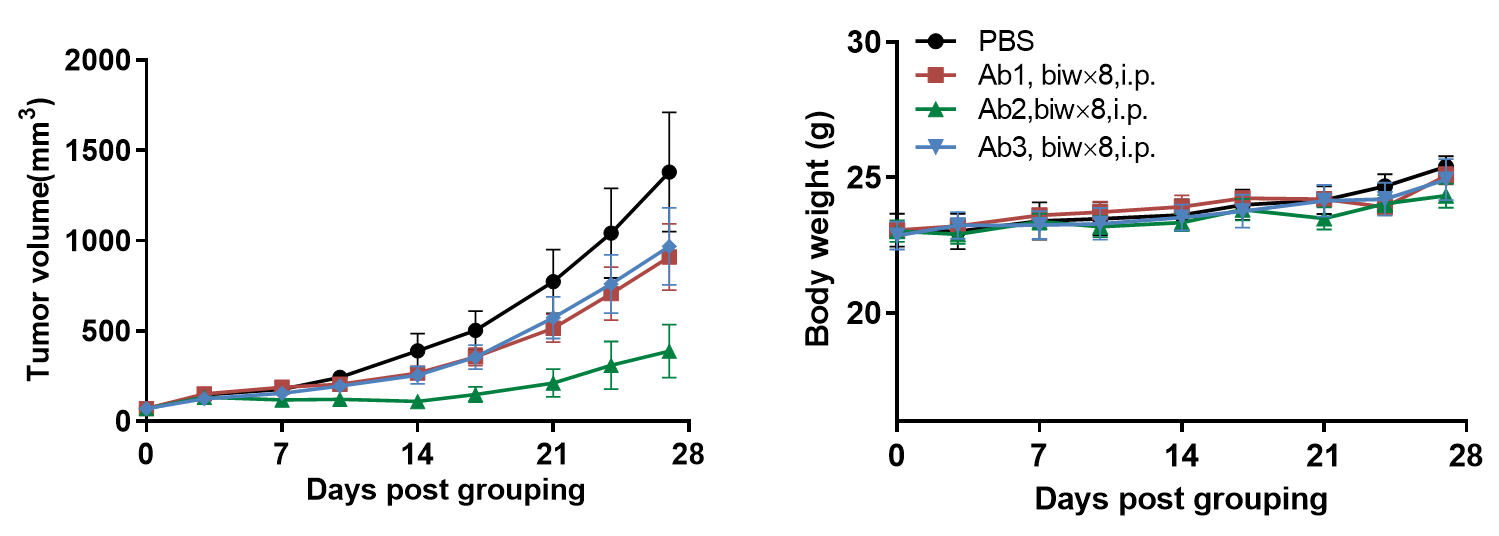
抗人CD16A抗体在B-hCD16A小鼠中的抗肿瘤活性。 (A) 抗人CD16A抗体抑制了B-hCD16A小鼠中MC38肿瘤的生长。小鼠皮下植入小鼠结肠癌MC38细胞(13周龄,雌性,n=7)。当肿瘤体积约为70mm3时,将小鼠分组并接受抗人CD16A抗体治疗,治疗剂量和方案如图B所示。 (B) 治疗期间体重变化。如图A所示,抗人CD16A抗体在B-hCD16A小鼠中有效控制了肿瘤生长,证明B-hCD16A小鼠为抗人CD16A抗体的体内评价提供了强大的临床前模型。值以平均值±SEM表示。
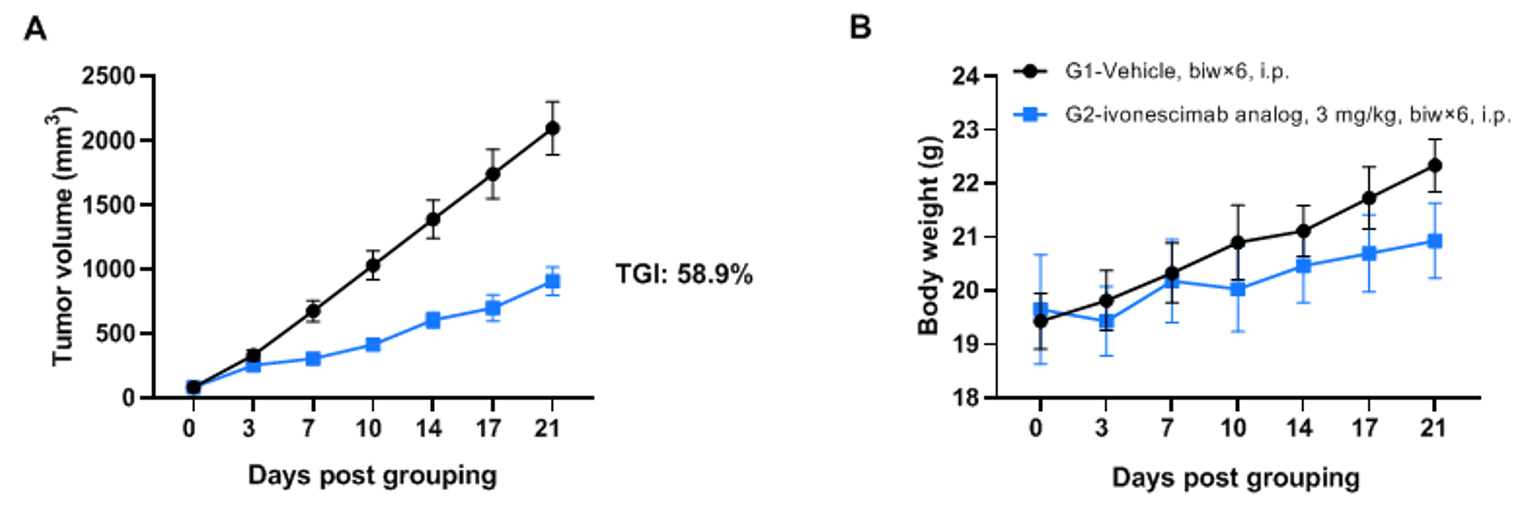
抗PD-1/VEGFA双特异性抗体(ivonescimab类似物,内部制备)在B-hPD-1/hPD-L1/hVEGFA小鼠中的抗肿瘤活性。 (A) 抗PD-1/VEGFA双特异性抗体抑制了B-hVEGFA MC38肿瘤在B-hPD-1/hPD-L1/hVEGFA小鼠中的生长。小鼠结肠癌B-hVEGFA MC38细胞皮下植入同源B-hPD-1/hPD-L1/hVEGFA小鼠中(8周龄,雌性,n=6)。当肿瘤体积达到约70-90mm3时,将小鼠分组并腹腔注射抗PD-1/VEGFA双特异性抗体。 (B) 治疗期间体重变化。如图A所示,抗PD-1/VEGFA双特异性抗体有效控制了B-hPD-1/hPD-L1/hVEGFA小鼠中的肿瘤生长,证明B-hPD-1/hPD-L1/hVEGFA小鼠为抗PD-1/VEGFA双特异性抗体的体内评价提供了强大的临床前模型。值以平均值±SEM表示。通过双向ANOVA检验确定显著性。*P < 0.05, **P < 0.01, ***P < 0.001。
| Category Name | Product Name | Product No. | Background | Action |
| T cell engager | B-hPD-1 mice | 110003 | C57BL/6 | |
| B-h4-1BB mice | 110004 | C57BL/6 | ||
| B-hCD3E mice | 110008 | C57BL/6 | ||
| B-hTIM3 mice | 110018 | C57BL/6 | ||
| B-hPD-1 mice plus | 110019 | C57BL/6N | ||
| B-dPD-1 mice | 110022 | C57BL/6 | ||
| B-hCD3EDG mice | 110039 | C57BL/6 | ||
| B-NDG mice | 110586 | NOD/SCID | ||
| B-NDG nude mice | 110587 | B-NDG | ||
| B-NDG hTHPO mice | 110590 | B-NDG | ||
| MORE >> | ||||
| Category Name | Product Name | Product No. | Background | Action |
| NK cell engager | B-NDG hIL15 mice | 110600 | B-NDG | |
| B-hCX3CL1 mice | 111257 | C57BL/6N | ||
| B-hB2M/HLA-E mice | 111352 | C57BL/6 | ||
| B-hB7-H3 mice (C) | 111806 | BALB/cCrSlcNifdc | ||
| B-hPD-1/hPD-L1/hCD94/hNKG2A mice | 111848 | C57BL/6 | ||
| B-NDG B2m plus/hIL15 mice | 111856 | B-NDG | ||
| B-hCTLA4/hTIGIT mice | 111860 | C57BL/6 | ||
| B-hHER2/hTROP2 mice | 111881 | C57BL/6 | ||
| B-hPD-1 plus/hTGFBR2 mice | 111890 | C57BL/6N | ||
| B-h4-1BB/hTIGIT mice | 112328 | C57BL/6 | ||
| MORE >> | ||||
| Category Name | Product Name | Product No. | Background | Action |
| Checkpoint-based BsAb | B-hPD-1 mice | 110003 | C57BL/6 | |
| B-h4-1BB mice | 110004 | C57BL/6 | ||
| B-hCD3E mice | 110008 | C57BL/6 | ||
| B-hTIM3 mice | 110018 | C57BL/6 | ||
| B-hPD-1 mice plus | 110019 | C57BL/6N | ||
| B-dPD-1 mice | 110022 | C57BL/6 | ||
| B-hCD3EDG mice | 110039 | C57BL/6 | ||
| B-NDG mice | 110586 | NOD/SCID | ||
| B-NDG nude mice | 110587 | B-NDG | ||
| B-NDG hTHPO mice | 110590 | B-NDG | ||
| MORE >> | ||||